原文來自 <Shrimp Book > 原作者Mathias Corteel,Hans J. Nauwynck
全文人工翻譯,相對理論的知識,避免枯燥,先上張讀書筆記-兩張圖看懂全文。
像其他節肢動物一樣,對蝦體表也具有一套非常有效的覆蓋物,該覆蓋物具有皮膚屏障和骨骼雙重作用,主要由外皮上皮細胞組成,通常該外骨骼也稱之為角質層。對于這層覆蓋物來說,將其理解為對蝦的動態器官是十分恰當且有必要的,特別是對于生長中的動物來說,外骨骼與周期性的蛻殼密切相關。為保證正常的生長,新的角質層一般“貯存”在舊角質層之下,一旦舊的褪去,新的柔軟角質層便伸展膨脹,動物體積也隨之增長。鑒于以上原因,對蝦大多生理機能都是伴隨著蛻殼節奏的不同而發生變化,它會在特定階段蓄積能量而用于快速生長。
目前關于十足類甲殼動物角質層的理解主要來源于鰲蝦下目和短尾下目,針對對蝦深入研究是缺乏的,例如對蝦角質層形態和組成方面并無詳細研究數據。因此,以下所述大多數內容是適用于所有甲殼動物的,針對對蝦的研究成果我們會給出明確說明。
形態學
甲殼動物外骨骼屬復雜的生物復合材料,主要由多聚糖、幾丁質、礦物質和一些脂肪組成,關于甲殼動物外表覆蓋物形態和生化組成的研究參見Compère等人綜述(2004),Fabritius等(2008)以大西洋龍蝦(Homarus americanus)為例闡述了節肢動物角質層分級組成結構。
生物高聚物幾丁質是外骨骼主要組成成分,它像鋼筋混凝土中的鋼筋一樣為角質層提供抵抗張力的特性,幾丁質基本單元是單糖N-乙酰-D-葡萄糖胺和D-葡萄糖胺,這些基本單元依靠“聚合劑”β-1,4-葡聚糖黏合成狹長的線性幾丁質。在節肢動物中,幾丁質是以反平行的方式排列:α-結晶形狀排列,18-25各幾丁質聚合鏈以直徑2-5nm的晶體為中心,這種幾丁質晶體核心構成中央軸,類似外套的蛋白質亞單元圍繞的中央軸周圍,這種幾丁質-蛋白質復合物構成了7.25nm寬,0.3μm長的微纖維。考慮到幾丁質和蛋白質最小組成單元大小,上述微纖維稱為納米纖維更符合實際。
蛋白質像混凝土一樣圍繞在幾丁質繩和納米纖維之間,這為角質層提供了不可滲透的特性,同時也增強了納米纖維抵抗機械擠壓的能力,上述蛋白組成和水分子多少都取決于角質層的機械特性(Skinner,1992;Andersen,1999)。一般蛋白組成包含兩類蛋白:分別是以共價鍵和非共價鍵形式與幾丁質或角質層其他成分結合的蛋白,蛋白質之間和蛋白質與幾丁質之間的共價結合可以使角質層變得堅硬,這一過程主要是由酚氧化酶催化的,它可以將苯酚分子轉化為可反應的醌類。分子通過共價鍵結合可以形成網橋,后者為角質層提供了深褐色或光滑的特性,這樣蛋白質就不會溶解到水中。非共價鍵結合蛋白屬自由蛋白,它只能與角質層其他成分以靜電或水和鍵形式結合,這些蛋白均可以很容易地被水溶性緩沖液提取。在甲殼動物中,蛻殼之后角質層外部及其真皮層均變得光滑。
與其他研究深入的短尾亞目甲殼動物相比,對蝦角質層干重中無機礦物質占到了30-50%,這使得礦化作用在針對對蝦角質層操作時作用不大。對蝦角質層礦物鹽主要是鈣、鎂和碳酸鍶,上述物質在自然條件下主要以晶體方解石形式存在,顯然,角質層的礦化增加的對蝦外殼的硬度。
電子顯微鏡分析表明幾丁質-蛋白質微纖維成打成束組成粗纖維,后者直徑可以達到100nm,角質層位置和功能的不同導致粗纖維排列也是不同的,因為后者很大程度上決定了角質層的最終機械特性。在大多數角質層中,粗纖維是眾多水平面層面組成的,水平面與角質層表面平行;在每一水平平面中,粗纖維沿著長軸貯存,每一長軸都平行于其他水平片段,不同片段的夾角會有一定度數變化,但角質層整體是一個螺旋形,類似夾板的扭曲結構,由不同水平分布的粗纖維疊加形成。從外表上看,垂直層面角質層類似晶片的結構是顯而易見的,側面看還具有拋物線特征。兩層相同方向的粗纖維之間的距離由薄片填充,在最外層粗纖維與其他層的纖維可能具有180°的夾角。
光學纖維鏡觀察發現完整的對蝦角質層分為四層,從外到依次為:上表皮、外表皮、內表皮和膜層,上表皮和和外表皮形成于蛻殼前期,內表皮和膜層形成于蛻殼后期,外表皮和內表皮中可以清晰地看到纖維小板。上表皮層在組成和結構均和其他層不同,該層非常薄,富含脂質、蛋白質和礦物質,但是不含幾丁質,它是機體抵抗外界因素的第一道屏障,主要參與機體滲透壓調節。外表層在蛻殼之前已經形成,蛻殼之后很短時間開始礦化,顏色變深,該層是整個外骨骼的主要支撐層。內表層與外表層之間具有明顯區別度,前者富含鈣,它可以很好的為外表層起支撐作用。在對蝦中,外表層較內表層具有更多的纖維小板,纖維小板之間的堆疊高度也要大于內表層,這點在蟹和龍蝦中是反過來的。膜層位于上皮細胞之上,是內表皮層分泌形成的最后一部分,該層一般不含礦物質,屬無鈣化層,含有的纖維小板也較薄,這些結構在蛻殼過程匯總具有重要的功能。
上皮細胞 層位于角質層最底部,主要由單層假復層上皮細胞組成,主要分泌產生整個外骨骼,包含許多精細的結構。靠近細胞層基部存在著星形色素細胞,此類細胞細胞質中色素可以視情況發生分布的改變,進而影響對蝦體色(Noël, 1994)。上皮細胞層還存在著其他類型細胞,如毛源性細胞,它可以釋放感覺剛毛;皮膚腺細胞,它可以通過角質層中的導管存在和釋放外分性物質;免疫佐細胞雖然主要還沒有研究透徹,但其功能被認為和昆蟲中的絳色細胞類似,主要參與角質層物質的合成(Locke,1994)。
圖1 15g重凡納濱對蝦角質層和腹足上皮細胞光學纖維圖片(蘇木精-曙紅染色)。ep:上表皮;ex:外表皮;en:內表皮;ml:膜層;epi:上皮細胞;ct:結締組織
蛻殼周期:覆蓋物形態和生理的周期性變化
甲殼動物蛻殼過程通常被描述為蛻殼周期,一般將舊殼蛻去到下一次蛻殼分為三個主要階段,按時間先后依次為:蛻殼后期、蛻殼間期和蛻殼前期,第一個針對甲殼動物蛻殼研究的學者采用字母對各個階段進行了標識,分別為A為蛻殼后期初、B為蛻殼后期末、C為蛻殼間期、D為蛻殼前期(Drach,1999)。研究初期,各個階段的區分主要是基于骨骼和相應組織的硬度,隨著附肢顯微鏡檢技術的發展,利用剛毛形態差異即可清晰直觀的區分各個蛻殼階段(Stevenson,1972;Aiken,1973;Lyle,1983;Musgrove,2000;Gorokhova,2002)。
不同學者采用不同的評價標準對許多對蝦屬種類進行了蛻殼周期的研究,這些物種包括桃紅對蝦(Penaeus duorarum)、墨吉對蝦(Penaeus merguiensis)、中國對蝦(Penaeus setiferus )、細腳濱對蝦(Penaeus stylirostris)、凡納濱對蝦(Penaeus vannamei)和斑節對蝦(Penaeus monodon)。目前,用于評判對蝦蛻殼階段的關鍵指標還是上皮細胞和剛毛的形態特征,包括色素沉淀程度、新剛毛的形成、剛毛內基質、剛毛腔內空腔的存在以及所謂的剛毛器官的形成(節瘤存在)。
表1. 關于對蝦蛻殼各階段特征的研究
| | | | | | |
| |
中國對蝦 細腳濱對蝦 | | | | | | | |
| |
| |
|
凡納濱對蝦 墨吉對蝦 | | |
| 5-13 | | | |
| |
| | | | |
斑節對蝦 | | | |
| | | |
接下來我們將結合已有研究和我們自己的研究對對蝦蛻殼周期各階段進行闡述。與鰲蝦科和短尾下目甲殼動物相比,對蝦的蛻殼周期一般短于前者,這使我們可以用有限的數字標識主要蛻殼階段。蛻殼前期,對蝦覆蓋物形態和生理變化已經得到了很好的描述,但是不同的學者采用各主要階段行了了細分,并采用不同的子階段進行了標識,這些研究表3和圖2中均有闡述(Drach,1939;Skinner,1985;Compère et al., 2004)。
表2 對蝦蛻殼各主要階段特征分析
| |
| | | | | |
| | | | 剛毛基部無基質存在,色素細胞在剛毛瘤基部排成一條線 | | | | | |
| | | | | | | |
| 上皮顆粒細胞 | |
| | | | | | | | | | | | | | | | |
| | | | | | | | | | |
| | | | | | | | |
|
| | | | | | | | |
| | | | | | | | 空隙最大,上皮組織邊緣呈鋸齒狀且存在反射率較高很高的白層 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
表3 凡納濱對蝦和斑節對蝦蛻殼主要階段特征詳解(根據Corteel et al.文章整理)
| |
A 蛻殼后期前 |
|
剛毛腔充滿透明上皮細胞基質 |
剛毛無內腔出現 |
|
B 蛻殼后期末 | 上皮細胞從剛毛出開始回縮 |
剛毛基部仍然存在上皮細胞基質 |
剛毛內腔開始出現 |
|
C 蛻殼期 | 上皮層以直線形式排列在剛毛節瘤基部 |
剛毛腔內部和基部均無上皮細胞基質 |
剛毛內腔清晰可見 |
|
D1 蛻殼前期初 | 上皮層與角質層分離,半透明區域開始形成 |
上皮層開始形成新的剛毛,但是不可見 |
剛毛腔內無上皮層基質 |
剛毛內腔清晰可見 |
|
D2 蛻殼前期末 | 半透明區域清晰可見 |
可以觀察到新角質層 |
新剛毛可見,但是折疊位于上皮組織層 |
剛毛腔內無上皮層基質 |
剛毛內腔清晰可見 |
|
| |
蛻皮后期
蛻殼后期初“A”(~A1,A2)
對蝦蛻殼之后身體會吸收水分使其角質層膨脹,這個階段是生長、角質層更新、四肢修復的唯一關鍵時間,新形成的角質層只包括上皮層和外皮層,因為它們在蛻殼前已經分泌形成了,覆蓋物的其他層結構還未硬化和礦化,因此此時的蝦殼顯得軟、易彎和易碎。
該階段上皮細胞通過孔道延伸到角質層中,與其緊密接觸,前者會繼續分泌物質到角質層以形成內表皮。該過程發展到最后,上皮層會從角質層脫落,并且剛毛腔的上皮層基質也會消失。
對蝦角質層狀態對對蝦行為具有重要的影響,蛻殼A期時對蝦步足活動不便,它們會將時間基本用于在水體中游泳,這過程也不攝食也無法抵抗同類的攻擊蠶食。
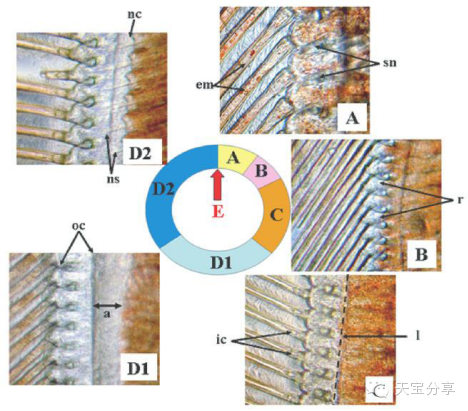
圖2 不同蛻殼階段凡納濱對蝦尾肢末梢顯微圖片,圖片以蛻殼先后順序順時針排列。A:蛻殼后期初,剛毛節瘤(sn)形成,剛毛腔內開始出現上皮層基質(放大倍數200X);B:蛻殼后期末,上皮層從剛毛處開始回縮(放大倍數100X);C:蛻殼間期,上皮層線性排列在剛毛節瘤基部,剛毛基部開始出現小內腔(放大倍數100X);D1:蛻殼前期初,舊角質層(oc)脫落導致其與上皮層之間空隙增大(放大倍數100X);D2:蛻殼前期末,上皮層形成新的折疊角質層(nc)和新剛毛(ns)(放大倍數100X);E:蛻殼期,舊殼脫落。
蛻殼后期末“B”(~B1,B2,B3,B4)
蛻殼進入B期后,上皮層和外皮層變硬,顏色也加深,整個外殼也隨之硬化和變深。內皮層也開始分泌形成,外皮層和內皮層礦化也在此時發生,上皮細胞開始變小并且夾雜在內皮層中。
與此同時,之前吸收的水分也逐漸被組織所取代,這一過程直到下一蛻殼階段開始時結束。外殼已經硬化,對蝦可以利用步足行動和攝食,之前所蛻之殼是其攝食的重要食物之一。
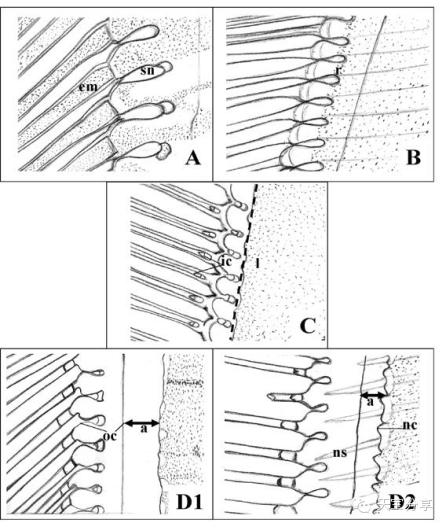
圖3 主要蛻殼階段對蝦尾肢末稍剛毛和上皮組織變化情況示意圖。A:蛻殼后期初,剛毛節瘤(sn)形成,剛毛腔內開始出現上皮層基質(em);B:蛻殼后期末,上皮層開始從剛毛處回縮(r);C:蛻殼期,上皮層線性(l)排列在剛毛節瘤基部,剛毛基部內腔(ic)清晰可見;D1,蛻殼前期初,舊殼脫落(a)導致其與上皮層之間出現空隙;D2,蛻殼前期末,上皮層開始形成新角質層(nc)和新剛毛(ns)。
蛻皮間期
蛻皮間期“C”(~C4)
通常認為蛻殼間期是對蝦蛻殼的休息期,該階段角質層完全形成,上皮細胞活動能力下降,形狀變類似骰子的立方形。該階段從生理角度考慮,對蝦會集中經歷蓄積能量,如糖原、脂質、鈣等,主要蓄積在肝胰臟和肌肉中。
蛻皮前期
蛻殼前期初“D1”(~D0,D1’,D1’’,D1’’’)
促進對蝦新角質層形成的荷爾蒙的分泌意味著蛻殼前期的開始,可觀察到的特征是上皮細胞開始延伸變長,衍化為具有很強機械運動能力的分泌細胞,同時,細胞核分裂能力也顯著增強且一直持續到蛻殼期。許多酶原(幾丁質酶和蛋白酶)被分泌到膜層,要在舊殼完全褪去之前分解吸收部分舊角質層,并將其凝膠化。在D1期,通過倒置電子顯微鏡首先看到的新角質層形成的主要變化發生在細胞頂端膜位置,小斑塊出現在頂端膜位置后不久變聚集合并,最終形成新的上皮層。蛻皮過程是從上皮層和舊角質層之間的蛻殼縫隙開始的。
該階段,肝胰臟和肌肉代謝能力用以為新角質層的形成提供能量。
蛻殼前期末“D2”(~D2,D3,D4)
隨著體積和活力的增加,上皮細胞進入D2期后開始分泌形成外表層,外表層位于上表層之下,短鏈低聚型幾丁質和表皮蛋白以幾丁質蛋白形式被合成分泌,幾丁質微纖維在幾丁質合成酶的催化下合,然后纖維被轉移到上皮細胞外表層區域,這一些列動作會消耗肝胰臟和肌肉積累的能量,但是在蟹類等甲殼動物中并沒有發現為此而過度消耗能量的現象(Csear et al.,2006)。
在該階段,舊角質層的降解達到最大程度,相關酶在蛻殼前期開始的時候已經被分泌到膜層和內皮層靠近基部的位置,蛻殼前期初形成的蛻殼裂縫充滿了蛻皮液。從此刻開始,在新角質層變得不能滲透之前,機體開始對葡萄糖胺、鈣和氨基酸進行再吸收。
舊角質層的變薄給了對蝦特定破殼位置,即蛻皮線,在對蝦中蛻皮線位于頭胸甲處,縱向步足處延伸,在蛻殼期對蝦可以輕松的從舊殼逃離。
由于舊角質層變弱和新舊兩層角質層的存在,對蝦在這個階段是不攝食的,這種情況一致持續到蛻殼后期末(B期),在此期間對蝦的代謝完全靠之前貯存的營養物質支撐。
蛻皮期
“E”期
該期對蝦會進行一系列肌肉收縮行為以脫去舊的外骨骼,頭胸甲處及其與第一腹節之間位置都會發生顯著性膨脹,此處像一個“艙口”一樣,對蝦會該處開始蛻去舊殼。
蛻殼周期的激素調控
像所有甲殼動物一樣,對蝦蛻殼也主要受以下四種內分泌激素調節:多肽類激素、類固醇激素、萜類激素和生物胺,整個蛻殼過程主要受多肽類的蛻皮抑制激素和類固醇類的蛻皮激素調控,這類激素作用是相對立的。
蛻皮抑制激素(MIH)由位于眼柄X器官的內分泌細胞產生,蛻皮激素主要由Y器官產生,后者位于臂前波斯管上皮細胞產生。
血淋巴中的蛻皮抑制激素含量維持在高位時,對蝦蛻殼過程基本停止并長期停留在蛻殼間期,蛻殼抑制激素減少的同時血淋巴中的蛻皮激素含量開始增加,Blais(1994)研究發現凡納濱對蝦Y器官分泌的主要蛻皮激素為3-去氫蛻皮松(3DE),它可以在上皮細胞中代謝演變為20-羥基蛻皮激素,后者可以刺激細胞為蛻皮做準備。蛻殼前期初結束時,血淋巴中蛻皮激素含量會維持在高位,進入蛻殼前后末時由于上皮細胞分泌功能的增強,蛻殼激素含量達到最大值,隨之蛻殼行為的推進,蛻皮激素會在蛻殼前期結束時出現顯著下降,之后各個蛻皮階段期含量一直維持在較低水平。
甲殼動物糖激素(CHH)最初被歸為調節碳水化合物代謝的核心激素,但事實上,該激素和蛻皮抑制激素(MIH)、性腺抑制激素(GIH)和下頜器官抑制激素(MOIH)一樣,同屬于神經肽價家族成員。上述激素一起協作調節甲殼動物蛻殼、糖代謝、繁殖和滲透壓調節等生理反應,甲殼動物的竇腺復合體也能分泌糖代謝激素(CHH),后者通過影響MIH間接對對蝦蛻殼激素的分泌產生抑制作用。
眾所周知,另一種物質——倍半萜大甲基法尼酯——在甲殼動物蛻殼調控中也起著重要作用。在蟹類和龍蝦的研究中,許多多功能單分子物質的作用已經被研究的很清晰,但這些物質在對蝦蛻殼和性腺發育過程中的作用最近才開始著手研究,例如有一種與昆蟲保幼激素相關的物質最近研究發現在對蝦下頜器官中也能產生。除此之外,針對MOIH的研究發現,該激素主要在蛻殼前期分泌,具有促進固醇類激素分泌功能,但是MIH可以抑制其的分泌。
除此之外還有兩種神經激素也必須加以說明,一種是甲殼動物心肌活動蛋白(CCAP),雖然已經被很多學者研究,但其在甲殼動物蛻皮過程中的作用還不是很清楚;另外一種是粘液素,該物質在調節昆蟲角質層sclerotisation process中作用已經很好被研究,但是在甲殼動物中雖然被證實也是最近研究發現的事,具體作用還需要進一步研究(Wilcockson,2008)。
雖然關于對蝦蛻殼調節的主要物質已經為人們所知,但各物質具體功能及其之間交互作用顯然比目前機理模式所呈現的復雜。目前為止,關于蛻殼激素含量在蛻殼前期開始降低的推測到也沒有被證實(Nakatsuji,2004),相反有證據表明蛻皮前期末蛻皮激素甚至出現了增加(Chung,2005)。
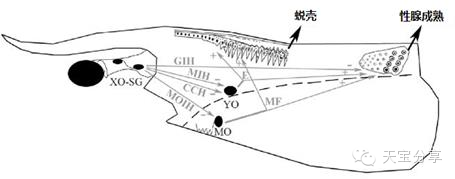
圖4 對蝦蛻殼內分泌調節原理圖。(XO-SG:X器官/竇腺復合體;GIH:性腺抑制激素;MIH:蛻皮抑制激素;CHH:甲殼動物糖代謝激素;MOIH:下頜器官抑制激素;YO:Y器官;E:蛻皮激素;MO:下頜器官;MF:甲基法尼酯)
在對蝦養殖過程中,由于蛻殼和和生長繁殖具有很好的相關性,因此養殖戶對蛻殼都比較重視。因此,基于多肽類激素GIH/MIH/CCH和蛻皮激素調控蛻殼過程的機理已經被很好的研究,性腺發育和肌肉生長一般是同步進行的,對兩者進行調控的關鍵器官是XO-SG。
在雌性對蝦親本養殖過程中有切除單側眼柄的習慣,養殖戶通過這一操作去除了內分泌激素對蛻殼的抑制。一般情況下,性腺抑制激素和蛻皮抑制激素對機體的抑制作用是相同步的,因此卵巢成熟和早蛻殼是同時出現的,幸存的XO-SG并不能替代已摘除眼柄的功能,因為蛻殼和卵巢的發育速度并不能及時降低。雌性對蝦的產卵時間和頻率都會增加,這會導致與卵黃形成和蛻殼相關的代謝行為過度旺盛,最終導致卵巢的崩潰(Palasios et al.,1993;Racotta et al.,2003)。雖然單側眼柄摘除并不能完全解決對蝦增殖中所有問題(Marsden et al.,2007),針對性腺抑制采取選擇性干預措施也不能很好的影響蛻殼和代謝,但是這些措施還是值得嘗試。





































 狗仔卡
狗仔卡



 發表于 2015-5-6 12:03:28
發表于 2015-5-6 12:03:28

 轉播
轉播 淘帖
淘帖 分享
分享 收藏
收藏 支持
支持 反對
反對 提升卡
提升卡 置頂卡
置頂卡 變色卡
變色卡 千斤頂
千斤頂 顯身卡
顯身卡 發表于 2015-5-16 18:07:53
發表于 2015-5-16 18:07:53






